")
")
Планктон
По определению (Horne, Goldman, 1994), "планктоном (название происходит от греческого слова "скиталец") являются пассивно плавающие или слабо передвигающиеся организмы, которые перемещаются по воле волн и течений". В состав планктона входят бактериопланктон - водные микробы, питающиеся растворенной и взвешенной органикой, фитопланктон - живущие в открытых водах водоросли, усваивающие углекислоту путем фотосинтеза, и зоопланктон - разнообразные беспозвоночные, питающиеся водорослями и бактериями. Особая роль в водных экосистемах принадлежит фитопланктону, так как он является первым звеном пищевой цепи и поставляет синтезируемое при фотосинтезе органическое вещество всем живущим в водоемах другим организмам.
Фитопланктон
Фитопланктон озер очень чувствителен к антропогенным воздействиям. В частности, в результате эвтрофикации озер происходит рост его биомассы и глубокая качественная перестройка видовых сообществ.
На рис. 3.1.1 показано, как изменялась биомасса фитопланктона в озере Бодензее в период с 1951 по 1994 г. в зависимости от концентрации растворенного фосфора. По мере накопления фосфора биомасса фитопланктона увеличивалась. Экосистема Бодензее оказалась весьма инерционной - рост биомассы продолжался в течение нескольких лет после того, как началось снижение концентраций фосфора в результате снижения фосфорной нагрузки, но затем, начиная с 1989 г., когда концентрация фосфора от максимального значения 80 мкг/л упала до 44 мкг/л, биомасса фитопланктона стала устойчиво снижаться.
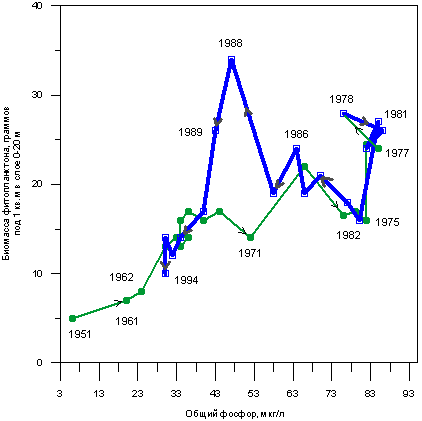
Рис. 3.1.1. Результаты наблюдений за концентьрацией фосфора и биомассой фитопланктона в озере Бодензее. С 1961 года наблюдения проводятся непрерывно, на графике приведены средние значения за каждый год. Kuemerrlin 1996.
![]() Загрузить исходные данные в формате Grapher 3
Загрузить исходные данные в формате Grapher 3
Данные об изменении биомассы фитопланктона в Байкале интерпретировать гораздо труднее, так как его концентрации здесь подвержены большой сезонной, годовой и пространственной изменчивости.
На рис. 3.1.2 показана динамика изменения биомассы фитопланктона в открытом Южном Байкале в период с 1948 по 1960 гг. по данным режимных наблюдений вблизи поселка Большие Коты. Из рисунка видно, что усредненная за год биомасса, определенная по найденной численности планктонных водорослей разных видов и объемам клеток этих водорослей, варьирует в разные годы в огромных пределах – от менее 10 до 290 граммов под 1 м2 в слое 50 м.
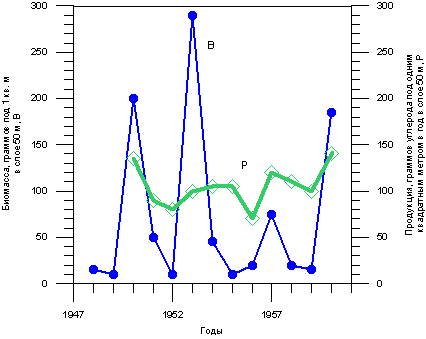
Рис. 3.1.2. Динамика биомассы и первичной продукции фитопланктона в Байкале. Kozhov 1963; Антипова 1963, цит. по Вотинцев и др., 1975, с. 145.
![]() Загрузить исходные данные в формате Grapher 3
Загрузить исходные данные в формате Grapher 3
В ходе режимных наблюдений определялась и первичная продукция фитопланктона, то есть то количество углекислоты, которое превращалось им при фотосинтезе в органическое вещество. Данные о первичной продукции также представлены на рис. 3.1.2. В отличие от биомассы, эта величина изменялась в значительно меньшем диапазоне, от 70 до 140 граммов углерода в год под 1 м2 в слое 50 м (105 ± 21 г/м2). Единственной причиной такого несоответствия мог быть недоучет каких-то значительных групп фитопланктона. О недоучете свидетельствуют и другие факты. Одним из них является уже упоминавшееся выше резкое снижение концентраций биогенных элементов летом 1948 г. в отсутствие выявленного потребителя: найденная при учете биомасса фитопланктона в тот год была очень мала (рис. 2.4.2). Второй косвенный признак - «продукционный парадокс» Байкала, описанный К.К. Вотинцевым и др. (1975), а именно характерные для Байкала отношения биомассы и продукции (Р/В) в диапазоне от 15 до 82, во много раз превышающие эти отношения, найденные для других озер (Р/В = 1-2).
{kind=link}
Перед подсчетом клеток фитопланктона различных видов его концентрируют путем пропускания через сита, либо путем отстаивания. При определении продукции радиоуглеродным методом клетки отделяют от воды с помощью ультрафильтров, которые удерживают даже мельчайшие клетки, в том числе пикопланктон. При «скляночном» методе определения первичной продукции, основанном на измерении концентрации кислорода, фильтрацию не производят вовсе. Если в ходе концентрирования пропусканием через сита или отстаиванием какие-то группы фитопланктона теряются, полученное отношение Р/В окажется завышенным. Именно это обстоятельство и может быть причиной байкальского продукционного парадокса. Отметим, что о надежности данных о первичной продукции свидетельствуют ее значения в диапазоне от 100 до 250 г углерода/м2 в год, полученные для временного интервала 1981-1985 гг. ( Бондаренко, Гусельникова, 1989).
По изложенным выше причинам найденная в разные годы биомасса фитопланктона варьировала в весьма широких пределах, и на основании данных о ней нельзя сделать выводов ни об эвтрофикации Байкала, ни об отсутствии эвтрофикации. Дополнительным осложнением, затрудняющим интерпретацию данных об изменении биомассы фитопланктона, является значительная пространственная изменчивость этого показателя. О такой изменчивости свидетельствуют данные непрерывной, выполняемой на ходу исследовательского судна регистрации хлорофилла (Гительзон и др., 1991; Левин и др., 1996). «Пятна» хлорофилла могут иметь размер от 1 до 30-50 км в поперечнике. Такой же характерный масштаб имеют неоднородности температуры, обнаруживаемые путем спутниковых наблюдений (Bolgrien et al., 1995; Семовский и др., 1998). Неоднородности перемещаются по Байкалу со скоростью нескольких километров в сутки.
Индикатором, гораздо более чувствительным, чем биомасса фитопланктона, является его видовой состав. Так, например, для Бодензее по данным долговременных режимных наблюдений известно (Kummerlin, 1996), что в период с 1896 до 1950 г. в составе фитопланктона постоянно доминировали диатомеи рода Cyclotella. С началом эвтрофикации в 1965 г. доминирующими стали диатомеи рода Stephanodiscus, которые уступили место диатомеям других родов лишь в 1992 - 1994 гг. В 1980 г. в больших количествах впервые появился самый грозный индикатор эвтрофикации - сине-зеленая водоросль Aphanizomenon flos-aquae, которая к настоящему времени вновь исчезла.
Изменения видового состава диатомовых водорослей нашли отражение и в осадочной летописи Бодензее (рис. 3.1.3). Упомянутые выше тенденции проявляются в этой летописи очень четко.
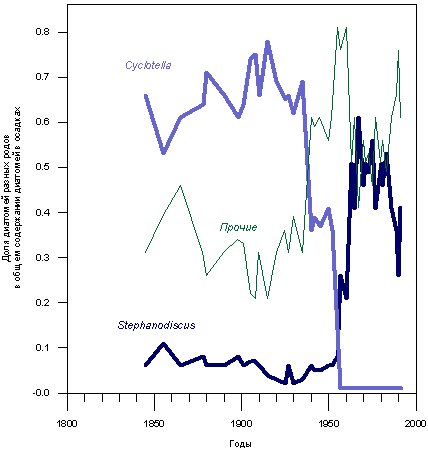
Рис. 3.1.3. Изменения видового состава створок диатомовых водорослей, захороненных в осадках озера Бодензее. Schroeder, 1996.
![]() Загрузить исходные данные в формате Grapher 3
Загрузить исходные данные в формате Grapher 3
Как уже указывалось выше, с конца 1940-х гг. в Южном Байкале в открытых водах на глубоководной станции в районе поселка Большие Коты специалистами Иркутского государственного университета проводятся регулярные режимные наблюдения за концентрациями различных видов фитопланктона. С середины 1960-х гг. до середины 1990-х гг. силами сотрудников Лимнологического института Сибирского отделения Российской академии наук проводился весенний учет фитопланктона на стандартной сетке разрезов по всей акватории Байкала. На основании данных этих измерений иногда делались выводы о том, что под действием антропогенных факторов уже произошли качественные изменения видового состава байкальского фитопланктона. Рассмотрим обоснованность подобных утверждений.
Очевидно, что для того, чтобы зарегистрировать качественные изменения видового состава, т. е. появление новых или исчезновение имевшихся ранее видов, достаточно было бы сравнить списки видов, составленные в разное время. Однако это совсем не простая задача для любой экосистемы, а в особенности для Байкала, отличающегося исключительно высоким уровнем эндемизма флоры и фауны. Благодаря развитию биологической науки в Байкале постоянно обнаруживаются новые, не зарегистрированные ранее виды планктонных водорослей. Но это отнюдь не означает, что их там не было раньше, возможно, что раньше по каким-то причинам они просто избежали внимания исследователей. Одной из причин обнаружения новых видов может быть усовершенствование методик сбора материала. Кроме того, постоянно происходит усовершенствование биологической номенклатуры и методов систематики, виды переименовываются, обнаруженные ранее - «закрываются», либо из состава одного вида выделяются еще один или несколько новых. Еще большую путаницу вносит включение в списки подвидов, разновидностей и форм. Наконец, иногда невозможно установить, появился ли «новый» вид в самом Байкале, либо случайно попал в открытые воды из мелководий или притоков. По указанным причинам об изменении состояния озерной экосистемы может однозначно свидетельствовать лишь устойчивое изменение состава наиболее типичных и массовых видов фитопланктона.
Исследования видового состава фитопланктона озера Байкал начаты еще в конце ХIХ - начале ХХ столетия. Основополагающий вклад в исследование качественного видового состава фитопланктона озера Байкал внес К.И. Мейер. Изучив образцы, собранные в 1916-1929 гг., он разделил Байкал по составу фитопланктона на 8 районов - открытый Байкал, пролив Ольхонские ворота, пролив Малое море, Баргузинский и Чивыркуйский заливы, Северная конечность Байкала, Мелководные заливы («соры») - Посольский, Истокский и Нижнеангарский, залив Провал; Селенгинское мелководье. По результатам своих исследований он опубликовал подробный список видов (Мейер, 1930).
По итогам дальнейших многолетних исследований фитопланктона различных районов Байкала в 50-х гг. вышла сводная работа В.Н. Яснитского и А.П. Скабичевского «Фитопланктон Байкала» (1957). В статье приводится систематический список, включающий более 120 видов и разновидностей водорослей. Указывается, что только 40 из них действительно живут и развиваются в открытом Байкале, остальные являются случайными в озере, они занесены из рек и соров. Из числа включенных в список видов 20 эти авторы считали типичными для открытого Байкала.
В 1959 г. список видов байкальского фитопланктона, включающий 99 видов планктонных водорослей и сопровождаемый кратким описанием биологии наиболее распространенных форм, был опубликован О.М. Кожовой (Кожова 1959).
Значительный вклад в изучение фитопланктона был внесен Н.Л. Антиповой. Ею подробно излагались сведения о периодических изменениях в составе фитопланктона и колебаниях численности доминирующих форм (Антипова, 1963). Н.Л. Антипова впервые для Байкала открыла цветение в подледный период перидиниевых водорослей и описала новые эндемичные виды беспанцирных перидиней (Антипова, 1955, 1963). Она опубликовала список планктонных байкальских водорослей, включающий 50 видов, из которых 20 были отмечены как принадлежащие байкальскому комплексу (Антипова, 1963). Л.Р. Изместьева и О.М. Кожова (Долгосрочное.., 1988) представили список из 70 видов и подвидов, обнаруженных в открытом Южном Байкале; из этого числа 12 видов были отмечены как наиболее массовые. Г.Ф. Загоренко и Г.С. Каплина (1988) опубликовали для той же станции вблизи поселка Большие Коты список, включающий 78 видов, из них 16 были названы доминирующими.
Г.И. Поповская (Вотинцев и др., 1975) по данным многолетних исследований всей пелагиали Байкала опубликовала список планктонных водорослей, включающий 92 вида. Из этого числа 30 отмечены ею как постоянные компоненты комплекса открытых вод, а 11-12 упомянуты как доминирующие.
В таблице 3.1.1 приведены списки пелагических планктонных водорослей открытого Байкала. В четвертой графе таблицы в качестве базового приведен список видов, типичных для открытого Байкала по мнению Г.И. Поповской (Вотинцев и др,. 1975). Список приводится здесь с небольшими изменениями, в основном связанными с изменениями номенклатуры. Из других колонок таблицы мы можем узнать, упоминались ли эти виды К.И. Мейером (1930), В.Н. Яснитским и А.П. Скабичевским (1957), О.М. Кожовой (1959) и Л.Р. Изместьевой и О.М. Кожовой (Долгосрочное.., 1988). Настоящий обзор в отношении видового состава байкальских планктонных водорослей не претендует на полноту. Несколько более подробные сведения на эту тему приведены в обзоре М.А. Грачева и Е.В. Лихошвай (1996), выполненном в соответствии с проектом ТACIS-BISTRO No.BIS/96/125/036. Однако данные, приведенные в табл. 3.1.1., позволяют предполагать, что пополнение списков происходило не в результате изменений в экосистеме Байкала, а вследствие развития биологической науки. Интересным примером является диатомея Nitzshia aciсularis, большие количества которой, наблюдавшиеся в 1970-1990 гг., по мнению Г.И. Поповской и др. (1997) и О.М. Кожовой и др. (1998) могут быть свидетельством антропогенных изменений. Однако этот вид был обнаружен в открытом Южном Байкале в значительных количествах еще К.И. Мейером (1930) и включен им в список (см. табл. 3.1.1) и потому не может считаться индикатором качественных изменений в экосистеме озера Байкал в результате промышленной революции.
| Наименования таксонов | Мейер 1930 | Яснитский, Скабичевский 1957 | Кожова 1959 | Поповская (Вотинцев и др. 1975) | Изместьева, Кожова (Долгосрочное... 1988) | |
|---|---|---|---|---|---|---|
| CYANOPHYTA | ||||||
| 1 | Synechocystis limnetica Popovsk. [1] | + К 100 | + 50 | |||
| 2 | Anabaena lemmermannii P.Richt. | + | + К 100 | + | ||
| CHRYSOPHYTA | ||||||
| 3 | Mallomonas vannigera Asmund [2 ] | + К * | + 50 | |||
| 4 | Uroglena volvox Ehr. [3] | A | A | A | A К | A |
| 5 | Dynobrion cylindricum Imh. | A П | A Б | A 50 | A К 100 | A 50 |
| 6 | D. divergens Imh. | A П | A Б | A 50 | A К | A |
| 7 | Chrysosphaerella baicalensis Popovsk. [4] | + К | + | |||
| 8 | Chrysosphaera melosirae (K.Meyer) Bourelli [5] | A | A Б | A | A К | A |
| BACILLARIOPHYTA | ||||||
| 9 | Aulacoseira skvortzowii Edmond, Stoermer, Taylor [6] | A П | A Б | A 50 | A К 100 | A 50 |
| 10 | A. baicalensis (K.Meyer) Sim. [7] | A П | A Б | A 50 | A К 100 | A 50 |
| 11 | A. distans (Ehr.) Sim. [8] | A | A | |||
| 12 | A. lirata var. alpigena (Grun.) Krammer [8] | + К | + | |||
| 13 | Cyclotella baicalensis (K.Meyer) Skv. [9, 10] | A П | A Б | A | A К | A |
| 14 | C. minuta (Skv.) Antip. [11] | A П | A Б | A 50 | A К 100 | A 50 |
| 15 | Stephanodiscus skabitschevskyi Popovsk. [12] | + | + | |||
| 16 | S. binderanus (Kutz.) Krieger [13] | A П | A Б | A 50 | A К 100 | A 50 |
| 17 | S. hantzschii Grun. | + 50 | + | + 50 | ||
| 18 | Fragilaria crotonensis Kitt. | A | A | A | A | A |
| 19 | F. capucina Desm. | A П | A Б | A | A | A |
| 20 | Synedra ulna var. danica (Kutz.) Grun. | + Б | + | + | + | |
| 21 | S. acus Kutz. [14] | A П | A | A | A К | A |
| 22 | Asterionella formosa Hass. | + Б | + 50 | + К | + 50 | |
| 23 | A. gracillima (Hantzsch.) Heib. | A П | A | A | A | A |
| 24 | Nitzschia acicularis W. Sm. [15] | A | A | A 50 | A К 100 | A 50 |
| DYNOPHYTA | ||||||
| 25 | Gymnodinium baicalense Antip. [16] | + Б ** | + 50 | + К | + 50 | |
| 26 | G. baicalense var. minor Antip. [16] | + Б ** | + 50 | + К | + | |
| 27 | G. coeruleum Antip. [16] | + Б ** | + 50 | + К | + | |
| 28 | Peridinium baicalense Kiss. et Zwetkow [17] | + Б | + | + К | + | |
| 29 | Ceratium hirundinella (O.F.M.) Bergh. | A | A Б | A | A К | A |
| CRYPTOPHYTA | ||||||
| 30 | Chroomonas acuta Uterm. [18] | + 50 | ||||
| CHLOROPHYTA | ||||||
| 31 | Monoraphidium arcuatum (Korsch.) Hindak [19] | + К 100 | + 50 | |||
| 32 | M. contortum (Thur.) Komarkova-Legnerova [20] | A * | A 50 * | A | A | |
| 33 | M. irregulare (Smith) Komarkova-Legnerova [21] | + | + | |||
| 34 | Sphaerocystis sp. [22] | + | + | |||
| 35 | Botryococcus braunii Kutz. | + | + | + | ||
| ULOTRICHALES | ||||||
| 36 | Koliella longiseta (Vischer) Hind. [23] | + 50 |
Обозначения:
A - виды, включенные в байкальский список К.И. Мейером (1930) и затем включавшиеся в последующие списки.
+ - виды, включавшиеся в байкальский список, начиная с 1957 г. и позднее.
П - постоянные компоненты байкальского фитопланктона по К.И.Мейеру (1930).
Б - истинные элементы байкальских фитоценозов по В.Н.Ясницкому и А.П.Скабичевскому (1957).
50 - виды, встречающиеся в Байкале в концентрациях более 50 тыс. клеток в литре по О.М.Кожовой (1975) и Л.Изместьевой, О.М. Кожовой (1988).
К - постоянные компоненты байкальского планктонного комплекса по Г.И. Поповской (Вотинцев и др. 1975).
100 - виды, встречающиеся в Байкале в концентрациях более 100 тыс. клеток в литре по Г.И.Поповской.
* - дано только название вида, но не подвида и формы;
** - дано только название рода.
Примечания к Таблице 3.1.1.
[1] S. limnetica описана как новый вид Г.И.Поповской в 1968 г.
[2] M. vinnigera описана как новый вид И.М.Балоновым в 1980 г., до этого учитывалась как Mallomonas sp.
[3] Uroglena volvox описана как новый вид В.Н.Яснитским (1924).
[4] Chrysosphaerella baicalensis описана как новый вид Г.И.Поповской в 1971 г.
[5] Chrysosphaera melosirae (K. Meyer) Bourelli = Epychrisis melosirae (K.Meyer).
[6] A. skvortzowii раньше учитывалась в Байкале как A. islandica предположение о том, что она является другим видом, было высказано на том основании, что, в отличие от последней, она образует споры (Edlund et al. 1996).
[7] До 1979 г. называлась Melosira baicalensis.
[8] = Melosira distans var. alpigena Grun.
[9] Описана как новый вид Б.В.Скворцовым в 1937 г.
[10] Из состава вида недавно выделен новый вид C. ornata (Flower, 1993).
[11] Выделена из состава С. baicalensis Антиповой в 1956 г.
[12] Описана как новый вид Г.И.Поповской в 1966 г.
[13] = Melosira binderanus Kutz.
[14] Включает разновидности radians и delicatissima
[15] Наличие этого вида в списке К.И.Мейера нужно особо отметить, так как некоторые исследователи считают, что этот вид появился в Байкале лишь в 1960-е годы.
[16] Описана как новый вид Н.Л. Антиповой в 1955 г., наряду с G baicalense var. minor и G. coeruleum.
[17] Описана как новый вид И. Киселевым и Н. Цветковым в 1935 г.
[18] Найдена в Байкале в 1976 г. О.М.Кожовой.
[19] = Ankistrodesmus pseudomirabilis Korsch.
[20] Ankistrodesmus falcatus (Corda) Ralfs var. spiriliformis G.S.West.
[21] Ankistrodesmus angustus Bern.
[22] Видовое название пока не дано.
[23] До 1980 г. учитывалась вместе с Monoraphidium arcuatum.
Из существенных изменений в списках следует упомянуть появление в них пикопланктонной водоросли Synechocystis limnetica, открытой Г.И. Поповской, весьма массового вида, речь о котором пойдет ниже. Эта водоросль не была обнаружена ранее вследствие несовершенства методик. Вид Anabena lemmermannii, включенный по данным более ранних исследований в список О.М. Кожовой (1959), вряд ли можно рассматривать как индикатор происшедших антропогенных изменений, так как в 1950-е годы антропогенная нагрузка на озеро была небольшой. Важным дополнением к списку К.И. Мейера стали динофитовые водоросли, которые были включены в список В.Н. Яснитского и А.П. Скабичевского еще в 1957 г. и потому также не могут рассматриваться как индикатор происшедших антропогенных изменений.
20 марта 1992 г. в открытом Байкале на станции вблизи поселка Большие Коты была обнаружена в больших количествах диатомовая водоросль Achnanthes minutissima (Кожова, Кобанова, 1994). Этот факт многократно упоминался О.М. Кожовой (например, Кожова и др., 1998, с. 119) как свидетельство качественных изменений фитопланктона Южного Байкала. Однако эта водоросль не является планктонной - она входит главным образом в состав обрастаний, используя в качестве твердого субстрата высшую водную растительность. Кроме того, следует иметь в виду, что она была найдена в больших количествах лишь в течение одного-единственного дня за весь период многолетних наблюдений, ее появление могло быть вызвано случайными причинами. Заметим, что О.М. Кожова и Г.И. Кобанова (1994) упоминают о том, что пробы воды (взяты 20 марта 1992 г.) наряду с A. minutissima содержали значительное количество взвешенного вещества неизвестного происхождения. По нашему мнению, разовое нахождение высокой концентрации A. minutissima в пелагиали Байкала по указанным причинам не может считаться свидетельством изменений в экосистеме Байкала.
Итак, автору настоящего обзора представляется очевидным, что качественный видовой состав ведущих байкальских планктонных диатомей по сравнению с началом ХХ столетия в Байкале не изменился. Для того, чтобы зарегистрировать более тонкие изменения, если они имели место, необходимы более подробные дополнительные исторические исследования.
Большие усилия исследователей Байкала были направлены на то, чтобы изучить количественные изменения состава фитопланктона в разные годы и сезоны. В частности, для станции в районе Большие Коты получен огромный объем данных для промежутка времени в 50 лет. Однако данные этих и других режимных наблюдений крайне трудно использовать для практической оценки состояния экосистемы озера Байкал, так как от года к году численность водорослей разных видов байкальского комплекса, как оказалось, меняется в исключительно широких пределах. Например, как уже упоминалось выше, численность ведущей байкальской диатомеи Aulacoseira baicalensis в поверхностных водах меняется от 1000 клеток в литре в немелозирные годы до 400000 клеток/л в мелозирные годы (см. рис. 1.1). Столь же широкий размах колебаний численности характерен и для других планктонных водорослей байкальского комплекса. На рис. 3.1.4 показана динамика изменения численности некоторых ведущих байкальских диатомей на станции в районе поселка Большие Коты по данным Н.Л. Антиповой (Долгосрочное.., 1988) для промежутка времени между 1973 и 1981 гг. Можно видеть, что концентрации Аulacoseira skvortzowii и Synedra acus колебались в пределах от аналитического нуля до 250 000 клеток/л. Особенно велики колебания численности Nitzshia acicularis, диапазон которых составляет от аналитического нуля до 1,5 млн. клеток/л (следует однако иметь в виду, что эта водоросль в силу своих малых размеров не вносит большого вклада в биомассу). О продолжающемся в 1990-е гг. резком колебании численности различных видов планктонных байкальских водорослей свидетельствуют данные совместного российско-английского проекта GEOPASS (Flower et al., 1998).
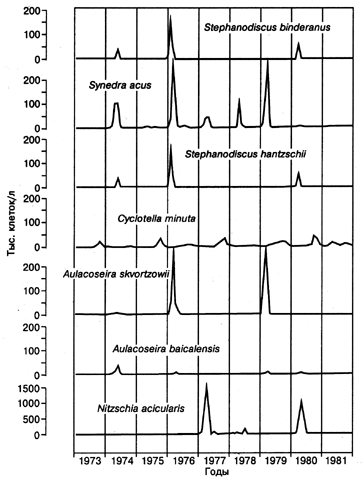
Рис. 3.1.4. Динамика изменения численности ведущих байкальских диатомовых водорослей в открытом Байкале на станции в районе пос. Большие Коты в 1973-81 гг. Данные Н.Л. Антиповой (Долгосрочное ...1988). Для пробоподготовки использовался отстойный метод (Utermoel 1936, 1958).
![]() Загрузить исходные данные в формате Grapher 3
Загрузить исходные данные в формате Grapher 3
Многочисленные попытки выявить корреляции видового состава диатомей с характеристиками климата и физико-химическими показателями, в том числе и те, которые предпринимаются в рамках проекта GEOPASS, пока не увенчались успехом.
Дополнительным осложнением является крайне неравномерное распределение различных видов фитопланктона по акватории Байкала, выявленное в работах Г.И. Поповской и др. (Вотинцев и др., 1975) и Н.А. Бондаренко и др. (Фактографическая.., 1997). На рис. 3.1.5 в качестве примера показано пространственное распределение Nitzshia acicularis, наблюдавшееся в мае-июне 1991 г. Можно отметить, что концентрации этой водоросли в поверхностных водах в это время варьировали в диапазоне от 1 тыс. до 350 тыс. клеток в литре. На фоне такой огромной изменчивости нельзя делать никаких выводов о том, что наблюдающаяся иногда высокая численность Nitzshia acicularis свидетельствует об антропогенных изменениях экосистемы Байкала. К сожалению, этого не понимают некоторые специалисты (Bondarenko, 1999).
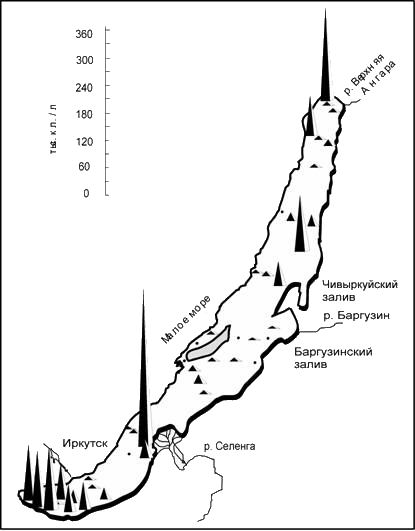
Рис. 3.1.5. Распределение численности Nitzschia acicularis в поверхностных водах Байкала. (Фактографическая база данных по фитопланктону оз. Байкал /Бондаренко Н.А., Воробьева С.С., Гусельникова Н.Е., Логачева Н.Ф., Помазкина Г.В., Лаврентьева Т.П. и Лобаева Л.А.; Правооблабатель - Лимнологический институт СО РАН; Свидетельство об официальной регистрации базы Данных № 97745 в РосАПО. Российская Федерация. Дата регистрации12.05.1997 г.).
Исключительно высокая вариабельность видового состава фитопланктона Байкала, по-видимому, определяется не только тем, что разные виды лучше всего приспособлены к физико-химическим условиям каждого данного момента времени, но и случайными причинами, например, наличием или отсутствием способных к размножению клеток в верхних слоях воды в начале вегетационного периода. Очевидно, что если такие материнские клетки в нужный момент времени отсутствуют, то этот вид в данном конкретном году не сможет доминировать, так как будет замещен тем или иным, быть может, даже менее приспособленным видом, если материнские клетки такого вида имелись в нужный момент времени в достаточном количестве.
Рассматривая вопросы конкуренции различных видов фитопланктона в Байкале, следует иметь в виду, что одним из главных участников этой борьбы за свет и биогенные элементы является пикопланктон. Как уже указывалось выше, эта группа водорослей была впервые обнаружена в Байкале Г.И. Поповской (Вотинцев и др., 1975).
Клетки пикопланктона имеют исключительно малый размер – от 1,5 до 3 мкм, не имеют кремнистого панциря и потому не захораниваются в осадках. Они крайне медленно оседают в поле тяжести и весьма существенно недоучитываются (как показали наши недавние опыты, приблизительно в 30 раз) при использовании для подсчета клеток фитопланктона, применявшегося при режимных наблюдениях отстойного метода Утермоля (Utermoehl, 1936; 1958). Однако они легко улавливаются ультрафильтрами при реализации радиоуглеродного метода определения первичной продукции (Кожова, Паутова, 1985; Бондаренко, Гусельникова, 1989), о чем уже сказано выше. Предположение о том, что именно пикопланктон в какой-то мере ответственен за «продукционный парадокс» Байкала, было высказано К.К. Вотинцевым и др. (1975), но не разрешило противоречия, так как найденный с учетом биомассы пикопланктона Р/В-коэффициент все равно остался очень высоким, поскольку величина В измерялась отстойным методом. Наконец, еще одно свидетельство в пользу большой роли пикопланктона в продукции Байкала было получено в российско-американском проекте (Beck et al., 1991), участниками которого было установлено, что 52-88% хлорофилла и 66-100% первичной продукции в Южном Байкале в сентябре-октябре 1989 г. приходилось на фракцию фитопланктона с диаметром клеток менее 10 мкм. Численность пикопланктона, измеренная в сентябре-октябре 1989 г. методом эпифлуоресцентной спектроскопии, достигала 10 тыс. клеток на мл (Boraas et al., 1991).
О весьма существенной роли пикопланктона как первого звена пищевой цепи в океанах стало известно лишь недавно. Роль этой группы в первичной продукции в пресноводных водоемах интенсивно изучается Известно, что концентрации пикопланктона в озерах иногда могут быть очень высокими (Ernst, 1990; Nagata 1990). В июле 1992 г. на Южном Байкале было впервые проведено комплексное исследование роли пикопланктона в рамках российско-японского международного проекта (Nagata et al., 1994). Оказалось, что максимальная численность пикопланктона достигает величин порядка 2х106 клеток/мл (2х109 клеток/л) в верхнем 15-метровом слое, что на три порядка больше, чем численность Aulacoseira baiсalensis в мелозирные годы. Пикопланктон отбирался на специальные окрашенные в черный цвет ультрафильтры и учитывался с помощью эпифлюоресцентного микроскопа по флюоресценции пигментов. При этом обнаружены две крупные группы пикопланктона: сине-зеленые водоросли, флюоресцирующие оранжевым цветом при возбуждении зеленым светом, и гораздо менее многочисленные зеленые водоросли, флюоресцирующие красным цветом при освещении голубым светом. Оказалось, что пикопланктону в указанный период принадлежала ведущая роль в первичной продукции (порядка 80%). Пикопланктон быстро потреблялся организмами, относящимися к размерному классу менее 20 мкм (жгутиковыми). Молекулярно-биологические и ультраструктурные исследования последних лет показали, что байкальский пикопланктон представлен не одним, а множеством видов (Семенова и др., 1998; Белых и др., 1999).
Продолжение количественных режимных наблюдений над видовым составом фитопланктона Байкала, несмотря на то, что из них пока трудно сделать практические выводы, представляет большой научный интерес, так как без результатов этих измерений мы не сможем понять ни важнейшие особенности механизма функционирования экосистемы озера, ни факторы поддержания биоразнообразия.
Первые детальные палеолимнологические исследования осадков Байкала, накопившихся в течение последнего столетия, были проведены в 1995 г. Flower et al. (1995; рис. 3.1.6).С помощью специального пробоотборника в Южном Байкале был взят короткий керн осадков с неповрежденным верхним слоем. Осадок был разрезан на слои толщиной от 2 до 5 мм и датирован свинцовым и цезиевым методами.
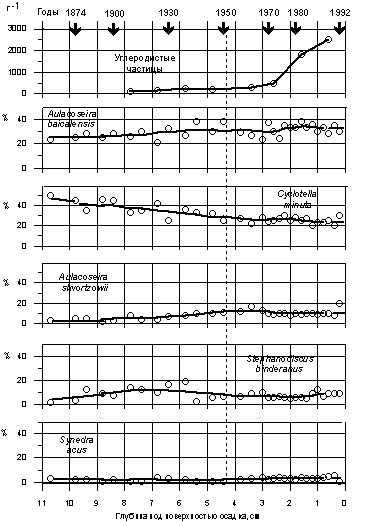
Рис. 3.1.6. Относительное содержание створок некоторых видов диатомовых водорослей (% от общей численности) и углеродистых частиц (шт. на грамм) в осадках озера Байкал разного возраста. Flower et al., 1995. Керн взят на глубине около 1400 м в Южном Байкале недалеко от истока Ангары.
![]() Загрузить исходные данные в формате Grapher 3
Загрузить исходные данные в формате Grapher 3
Промышленная революция в Сибири нашла свое отражение в байкальской осадочной летописи. Как видно из рис. 3.1.6, в период с 1950 до конца 1980-х гг. произошел резкий рост концентрации так называемых углеродистых частиц, мельчайших сферических частиц углерода, образующихся при сжигании угля при высокой температуре в топках крупных тепловых станций. Такие частицы не образуются при низкотемпературном сжигании, в частности, в паровозных топках. Можно видеть, что до начала 30-х гг. углеродистые частицы в осадках Байкала полностью отсутствовали.
Однако промышленная революция в Сибири, начавшаяся в 1950-е гг., никак не сказалась на видовом составе захороненных в осадке байкальских диатомей. На том же рис. 3.1.6 приведены данные об относительном содержании кремнистых створок пяти ведущих видов диатомей в осадках Байкала. Можно видеть, что в период с 1950 до 1992 г. содержание Aulacoseira baiсalensis составляло 30%, Cyclotella minuta - 25%, Aulacoseira skvortzowii - 10%, Stephanodiscus binderanus- 8 -10%, Synedra acus - 2 -3%. Контраст с осадочной летописью Бодензее (рис. 3.1.1) очевиден: в Байкале за те же последние 50 лет не изменился не только набор ведущих видов, но даже и их соотношения. Таким образом, палеолимнологические исследования не дали никаких свидетельств антропогенной эвтрофикации Байкала. Что же касается тех небольших изменений, которые произошли в период с середины прошлого столетия до 1950 г., то они, согласно мнению Flower et al. (1995), могли быть следствием небольшого потепления климата по окончании «малого ледникового периода».
В дальнейшем аналогичным исследованиям были подвергнуты короткие керны с неповрежденным верхним слоем, взятые в других 20 точках Байкала (Mackey et al.,1998). Авторы этого исследования делают следующий однозначный вывод: "наиболее молодые осадки, особенно те, которые взяты в южной котловине и на самом севере Байкала, содержат свидетельства антропогенных загрязнений в форме повышенных концентраций свинца и сферических углеродистых частиц, что, в частности, подтверждает заключение о преимущественном воздействии атмосферных источников загрязнения на южную котловину. Однако осадочная диатомовая летопись не дает никаких свидетельств изменения качества вод открытого Байкала вследствие поступления загрязнений через атмосферу и эвтрофикации" (Mackey et al., 1998, с. 1011).
Недостатком палеонтологического метода является то, что многие виды планктонных водорослей, в том числе и некоторые диатомеи, в осадках Байкала не захораниваются, а растворяются еще в водном теле. Этим объясняется, например, отсутствие в осадках Байкала весьма многочисленной диатомеи Nitzsсhia acicularis. Причина селективного растворения диатомей в водной толще до последнего времени была совершенно непонятна, так как, хотя створки некоторых видов исчезают целиком, степень сохранности даже мельчайших элементов ультраструктуры других водорослей, например, A. baicalensis, A. skvortzowii, С. minuta и др., остается очень хорошей. Недавно установлено, что причиной селективности растворения створок морских диатомей может быть то, что в этом процессе принимают участие специализированные микроорганизмы (Bidle, Azam, 1999): растворение кремнистых створок начинается лишь после того, как эти микроорганизмы разрушают выстилающие их поверхности органические макромолекулы.
Итак, накопленный огромный объем данных о байкальском фитопланктоне в целом говорит о том, что его состав за весь период научных исследований, начиная с первых десятилетий ХХ века, не претерпел существенных изменений. Этот вывод полностью соответствует рассмотренным в разделе 2 данным химических исследований, которые говорят о том, что причин для изменений фитопланктона не было, так как химический состав вод остался неизменным.
Зоопланктон
В состав зоопланктона Байкала входят жгутиковые, инфузории, коловратки и ракообразные. Наиболее важными представителями ракообразных является байкальский эндемичный рачок Epischura baicalensis и широко распространенный вид Cyclops kolensis. Характерный размер тела этих ракообразных - 1 мм. Кроме того, большое значение в Байкале имеет так называемый макрозоопланктон, представленный эндемичным рачком Macrohectopus branickii с характерным размером тела порядка 2-3 см. В настоящем обзоре мы рассматриваем только те аспекты состояния байкальского зоопланктона, которые могут характеризовать изменения состояния экосистемы озера в целом.
Имеющиеся данные не позволяют утверждать, что за время научных исследований Байкала произошли какие-либо качественные изменения состава его зоопланктона - в экосистеме озера не появился ни один новый вид, и ни один из видов не вымер. Об этом можно судить, в частности, по данным, приведенным в монографиях (Kozhov, 1963; Lake Baikal..., 1998).
Результаты режимных наблюдений за численностью зоопланктона как на станции в районе поселка Большие Коты, так и по всей акватории Байкала показали, что эта численность подвержена значительным сезонным, годовым и пространственным колебаниям. На рис. 3.1.7 показана динамика изменения биомассы планктонных ракообразных в открытом Байкале в верхнем 250-метровом слое на станции в районе поселка Большие Коты (Kozhov,1963). Отметим, что размах колебаний биомассы планктонных ракообразных велик, но существенно меньше, чем тот, который характерен для фитопланктона. В частности, концентрация планктонных ракообразных никогда не падает до нуля. Попытки выявить какие-либо четкие корреляции колебаний биомассы зоопланктона с колебаниями биомассы фитопланктона по результатам наблюдений 1946 - 1961 гг. не увенчались успехом. Установлено, что зоопланктон более активно размножается в те периоды, когда температура поверхностных вод повышена. Максимальное развитие Epischura baicalensis и Cyclops kolensis имеет место в разные годы, что является, по-видимому, свидетельством их конкурентных пищевых взаимоотношений.
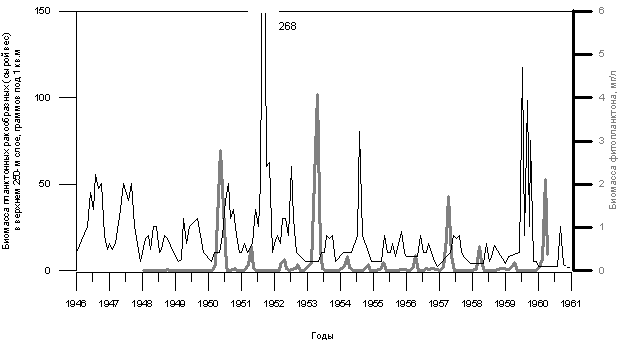
Рис. 3.1.7. Динамика изменения биомассы планктонных ракообразных (тонкая черная линия) и биомассы фитопланктона (толстая серая линия). Кozhov 1963, pp.235, 237, 238.
На рис. 3.1.8 показана динамика изменения биомассы зоопланктона в Байкале в период с 1961 - 1985 гг. (Афанасьева, Игнатов, 1992). Приведенные значения являются усредненными величинами, полученными при съемках по всей акватории Байкала. Э.Л. Афанасьева и А.В. Игнатов считают, что эти данные статистически достоверно указывают на снижение биомассы зоопланктона в Байкале в период с 1969 по 1985 гг. по сравнению с периодом 1961 - 1968 гг. приблизительно в 2 раза. Эти авторы отмечают, что причиной снижения биомассы зоопланктона могло быть влияние нескольких факторов. Например, на период с 1969 по 1974 гг. был введен временный запрет на промышленный лов байкальского омуля, в середине 1960-х гг. произошло резкое сокращение численности пелагического бычка-желтокрылки и др.
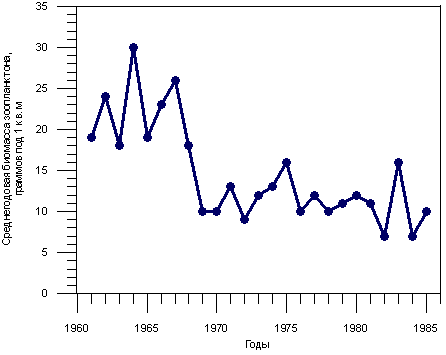
Рис. 3.1.8. Динамика изменения биомассы зоопланктона в Байкале в 1961 - 85 гг. Афанасьева, Игнатов, 1992.
![]() Загрузить исходные данные в формате Grapher 3
Загрузить исходные данные в формате Grapher 3
В то же время О.М. Кожова и А.М. Бейм (1993), проанализировав свою, весьма представительную выборку наблюдений, утверждают, что биомасса зоопланктона в Байкале в указанный период не снижалась.
В средствах массовой информации в 1970 - 1980 гг. неоднократно появлялись сообщения о массовой гибели эндемичного байкальского планктонного рачка Epischura baicalensis, якобы происшедшей по причине загрязнения вод Байкала стоками Байкальского целлюлозно-бумажного комбината. При этом журналисты не принимали во внимание, что существуют причины, вызывающие естественную смертность рачков. В частности, срок жизни этого организма равен одному году (Afanasyeva, 1998). Кроме того, известно (Кожова, Бейм, 1993), что Epischura baicalensis время от времени поражается грибком Saprolegnia, и этот процесс иногда охватывает значительные части акватории, в особенности в теплые годы.
Большой интерес представляет методика измерения распределения макрогектопуса в водах Байкала, основанная на использовании эхолокации и отлова сетями (Rudstam et al., 1992). К сожалению, эта методика пока не применена ни для всей акватории Байкала, ни для режимных многолетних наблюдений.
В предыдущие годы при режимных наблюдениях недостаточное внимание было уделено той группе зоопланктона размерного класса менее 20 мкм, которая питается пикопланктоном, (Nagata et al., 1994). В составе этой группы преобладали жгутиковые. Недостаточно изучены характеристики и других групп байкальского микрозоопланктона.
Автор настоящего обзора не нашел в опубликованной литературе каких-либо однозначных свидетельств того, что зоопланктон Байкала изменился под действием антропогенных нагрузок.
Бактериопланктон
Бактериопланктон Байкала пока изучен недостаточно. Во многом это объясняется тем, что байкальские бактерии, как и микроорганизмы из других водоемов, в том числе из мирового океана, крайне трудно культивируются в лаборатории, а потому не могут быть надежно идентифицированы классическими микробиологическими методами. Новые возможности для исследования байкальского бактериопланктона появились благодаря развитию методов молекулярной биологии (Беликов и др. 1996; Белькова и др. 1996), однако эти методы пока не внедрены в практику режимных наблюдений.
Имеющиеся сведения (см. например, Кожова, Бейм, 1993) о бактериопланктоне открытого Байкала не позволяют говорить о том, что он изменился под действием антропогенных нагрузок. Однако известно, что вблизи расположенных на берегу Байкала поселков и в устьях впадающих в него рек поверхностные воды Байкала иногда существенно загрязнены кишечной палочкой (E. coli), источником которой являются фекалии человека и животных. Кишечную палочку нельзя рассматривать как компонент байкальского бактериопланктона, так как в условиях водоема она не размножается. Данные об этом загрязнении будут рассмотрены в разделе о качестве байкальской воды.
Перепечатка без согласия автора запрещена. E-mail для контактов: grachev@lin.irk.ru
Публикация книги в сети интернет выполнена по гранту РФФИ-байкал №05-07-97200 E-mail: jonin@lin.irk.ru